Uno de los problemas transcendentales que tenemos que afrontar, es el hecho de que la supervivencia de las plantas se ve afectada por el calentamiento global. Las plantas experimentan estrés fisiológico debido a olas de calor, sequías, incendios o heladas; cuando son de gran magnitud y duración las conducen al deceso. Pero, ciertas plantas evitan su declive reparando el daño y recuperándose parcial o completamente. La recuperación post-estrés se realiza a varias escalas: a nivel de hoja, árbol y hasta de paisaje. Para planear su protección y recuperación, este grupo de investigación se da a la tarea de comprender el comportamiento del flujo del agua a través de su arquitectura hidráulica (xilema) bajo condiciones extremas (ver Fig. 1 y 2). Actualmente, no se tiene una computadora para modelar la arquitectura hidráulica de un árbol desde la raíz a las hojas, es por ello, que los estudios se enfocan en una de sus estructuras hidráulicas primordiales.
![Figura 1. Arquitectura hidráulica del árbol. a) Corte transversal de una rama en donde se observan sus estructuras internas (Anacauita); b) imagen de Traqueidas y vasos del Xilema por MEB [2]](/es-mx/AlmacenDigital/Gaceta/Gaceta_Mayo_Junio_2022/PublishingImages/02_analisis-formacion-vapor-conifera_01.jpg)
Figura 1. Arquitectura hidráulica del árbol. a) Corte transversal de una rama en donde se observan sus estructuras internas (Anacauita); b) imagen de Traqueidas y vasos del Xilema por MEB [2]
Así, se estudia y analiza mediante modelado numérico el flujo de la sabia a través de las fosas de un pino, especie Picea mariana, y se emula el proceso de la formación de la cavitación en sus conductos para conocer el umbral que produce su embolia generalizada (muerte), con el fin de evitar dichos valores mediante los cuidados de las plantas y su entorno.
Actualmente, en la literatura científica se asume que el agua es impulsada a través de los conductos del xilema por gradientes de tensión inducidos por la pérdida de agua (evaporación) que sucede en las hojas a través de los estomas (transpiración) formando columnas continuas de agua desde las raíces hasta los órganos foliares. Esta hipótesis se denomina teoría de la cohesión-tensión (CTT) y fue formulada por Dixon y Joly (1895)
[1] y renovada por Tyree y M. H. Zimmermann en 2002
[2]. Esta teoría sienta las bases de la existencia de la presión hidrostática “negativa". Se considera que la arquitectura hidráulica del xilema es una "tubería" aislada de todos los demás sistemas y que el flujo que sostiene por definición es inestable; ya que la tracción del agua al ser negativa (metaestable) es fácilmente quebrantada lo que provoca cavidades.
Esto nos revela que los árboles coexisten con la cavitación y tienen mecanismos alternativos para recuperarse, evadirla o tolerarla, hasta cierto punto
[3]. Dichos mecanismos ocupan tiempo, a veces de días a semanas
[4], de meses a años
[5] o incluso décadas[6]. Se conocen tres procesos de formación de embolismo:
i) Embolismo inducido por sequía, la tensión del agua en el xilema se vuelve tan alta que el aire disuelto se expande en los vasos y/o traqueidas obstruyendo la conducción del agua; ii) Embolismo inducido por congelación invernal, durante la congelación las burbujas de aire son expulsadas de la solución formando conglomerados, éstos a su vez, forman núcleos gaseosos (burbujas) que al arribar la descongelación, se expanden formando embolias locales; iii) Embolismo inducido por patógenos.
Hay especies que recargan su xilema y recuperan su función hidráulica, a la fecha, se desconocen los detalles finos de tal recuperación. Zeppel
et al.
[7] encuentran que las especies de zonas áridas tienen mayor recuperación que las especies de zona mésica. En la Fig. 2 se resume en forma pictórica lo que sucede en un árbol funcional y los mecanismos que se reconocen para escapar, tolerar o evadir la sequía. Además, el sistema arquitectónico hidráulico de los árboles es redundante.
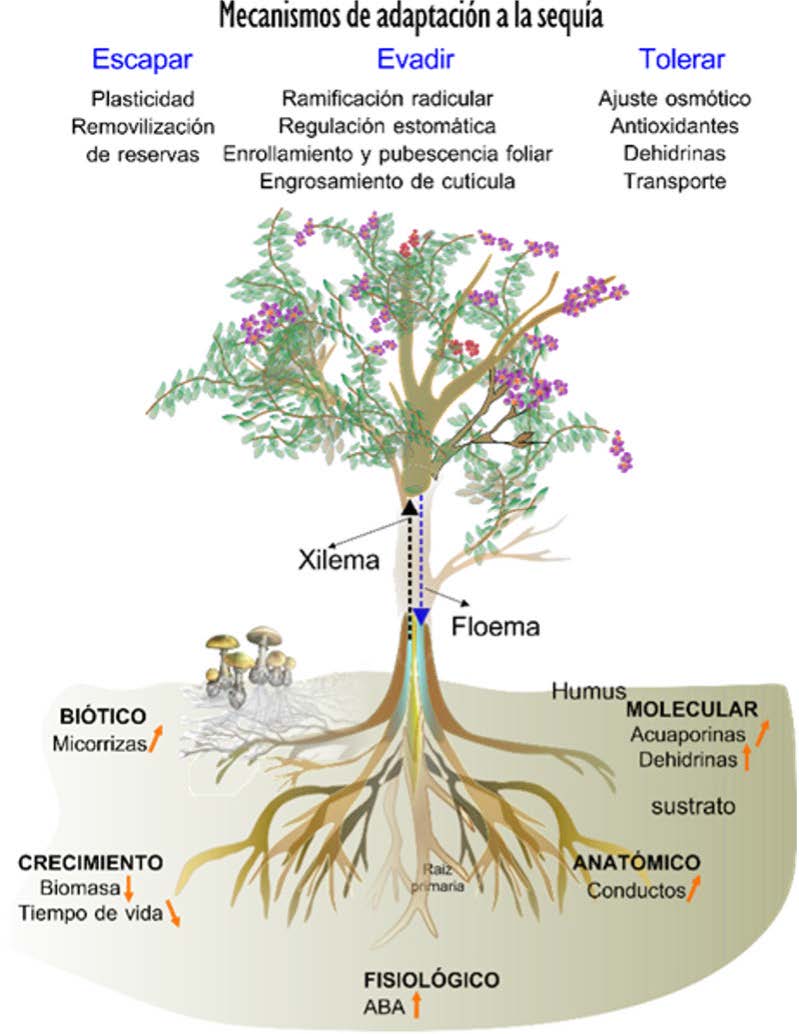
Figura 2. a) Vista global de los procesos que suceden en una planta vascular. El xilema realiza la transferencia de agua del suelo a la atmósfera. El floema transporta azucares y otros metabolitos desde las hojas a los tejidos. En la sección inferior se indican los mecanismos de resistencia a la sequía ya sea evitándola, resistiéndola o tolerándola. Efecto positivo, negativo y tendencia
Sistema del tejido vascular: Xilema
El xilema se compone principalmente de dos elementos por donde fluye el agua: las traqueidas (alargadas y arregladas en filas verticales) y vasos (con cribas en sus extremos), ambas son conductos formados por células muertas especializados para conducir el agua desde la raíz. Su longitud depende de la especie y de las condiciones ambientales. Las traqueidas alcanzan una longitud < a 5 [mm] y diámetros ~ 30 [μm]. Los vasos alcanzan longitudes de metros y sus diámetros oscilan entre 20 y 70 [μm]. Las paredes de los elementos se conectan transversalmente mediante válvulas llamadas “fosas o punteaduras" cuyos diámetros se encuentran en el intervalo de 9 a 18 [μm] (ver Fig. 1b).
Para emular el flujo ascendente a través de las traqueidas se considera la geometría de una fosa aureolada incrustada en la pared interfacial (ver Fig. 3a). El flujo axial se desvía hacia la membrana perforada (Margo) debido a que tiene su centro sólido. Para nuestro caso, usamos la geometría de una fosa de pino (Picea mariana). La fosa es aureolada y se modela como un disco constituido por una placa interior sólida y un borde exterior perforado (Margo) (ver Figs. 3 y 4)
![Figura 3. (a) Imagen SEM de las fosas [2]; (b) modelo 3D [9]; (c) corte de una fosa. dirección del flujo vertical y horizontal respectivamente [12]](/es-mx/AlmacenDigital/Gaceta/Gaceta_Mayo_Junio_2022/PublishingImages/02_analisis-formacion-vapor-conifera_03.jpg)
Figura 3. (a) Imagen SEM de las fosas [2]; (b) modelo 3D [9]; (c) corte de una fosa. dirección del flujo vertical y horizontal respectivamente [12]
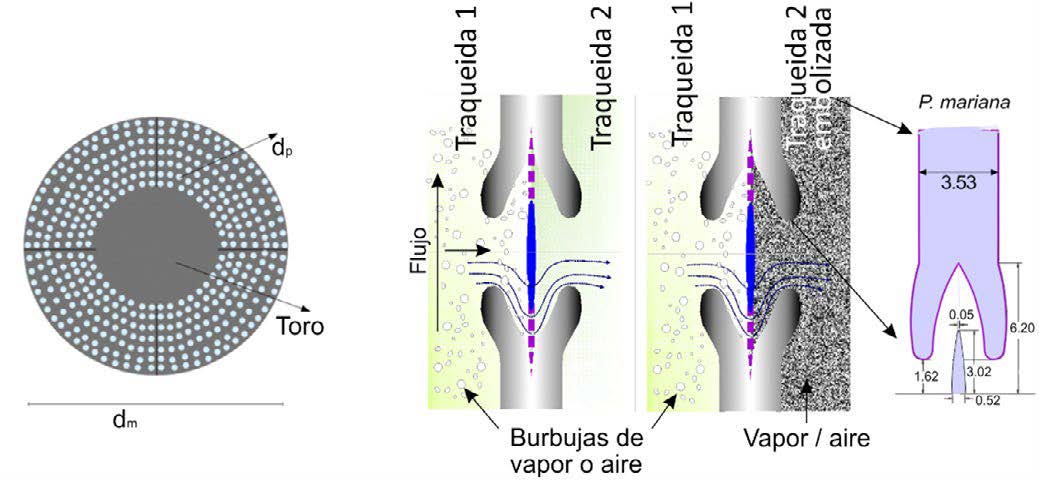
Figura 4. Geometría para el modelado numérico del flujo. a) Se emula la membrana de las fosas (margo) a una geometría de placa perforada que permite cambiar la resistencia del flujo variando tanto el diámetro de los orificios como su número; b) Trayectoria representativa del flujo longitudinal y transversal de agua entre dos traqueidas donde al pasar por las aberturas sufre una transición de fase de líquido a vapor; c) Dimensiones principales de la areola de la fosa y del margo
Resultados
Con el fin de estimar el rol de los componentes que integran la fosa, como los bordes, el toro y el margo, se construyen los modelos bajo las siguientes condiciones: velocidad de flujo en los conductos de 0.3 a 0.8 [mm s-1]
[2], un número de Reynolds* de alrededor de 0.08; por lo que las ecuaciones de gobierno se resuelven considerando flujo laminar**. A partir de un análisis de independencia de malla, se llega a un mallado tetraédrico de 5 millones de elementos con un refinamiento en la zona del margo de la fosa (ver Fig. 5).
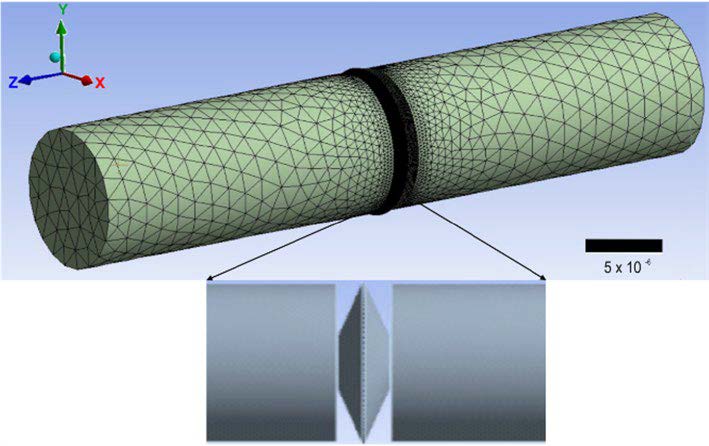
Figura 5. Mallado tetraédrico con 5 millones de elementos con refinamiento en la zona de la fosa para modelar el desplazamiento del flujo transversal entre traqueidas a través de una fosa
Las ecuaciones de continuidad y cantidad de movimiento se resuelven con el
software Ansys Fluent, utilizando el método de volúmenes finitos. El fluido de trabajo es agua con propiedades estándar y el flujo se considera en estado estacionario. Para resolver el acoplamiento de la presión con la velocidad se aplica el algoritmo SIMPLE (Semi-Implicit method for pressure linked equations). Se utiliza una discretización espacial de segundo orden para la presión y el momentum. Se fija el gasto másico de entrada a 6.25x10-11[kg s-1], y se impone una presión de salida en el orden de la presión de vapor del agua ~ 3325 [kPa].
Debido al tamaño de los elementos del xilema y a las propiedades hidrófobas del material; se selecciona la condición de deslizamiento en la pared (esfuerzo cortante nulo en la pared) por lo que no existe resistencia al flujo por las paredes.
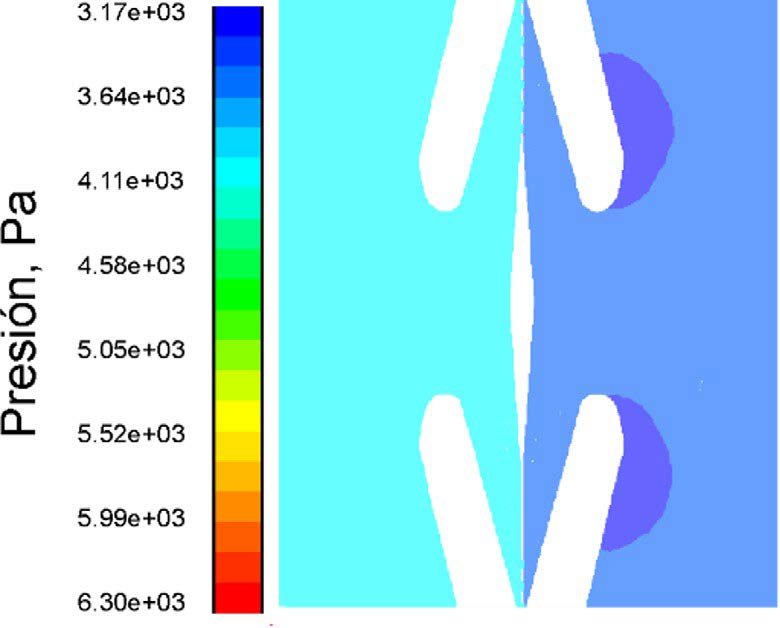
En general, la entrada de la fosa protegida por bordes de la aureola genera una región de alta velocidad de flujo (Figs. 3b y 6). Las Figs. 6a y b muestran una solución para la presión y la velocidad considerando la geometría de una fosa de la especie seleccionada. Se obtienen contornos similares del flujo variando tanto la velocidad de entrada como la presión. El flujo se acelera en la parte del margo cercana al toro.
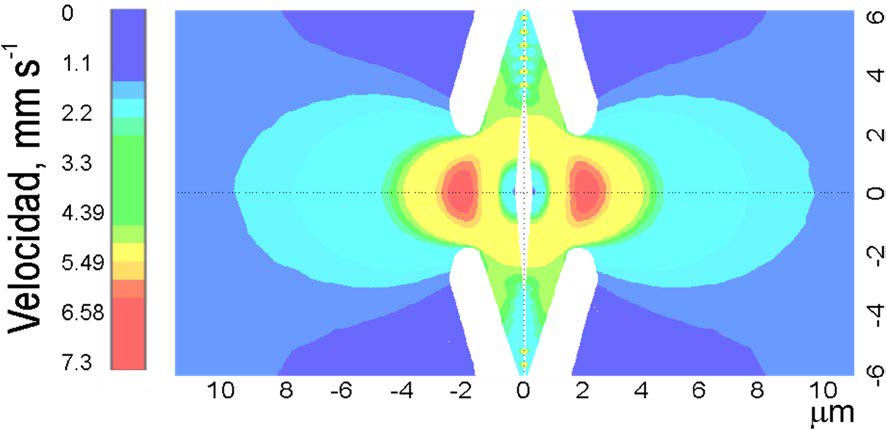
a)
b)
Figura 6. Resultados del modelo representativo mostrado perfiles de velocidad y presión del flujo a través de la cámara del foso. a) Iso-superficies de velocidad en [mm s-1], los números a la izquierda y borde inferior indican distancias en [μm]; b) Perfiles de presión en donde se aprecian zonas de baja presión en la traqueida 2
Para resolver el flujo bifásico se aplica el modelo de la mezcla considerando una fracción de vapor inicial de 0.001%. Se realizan diversas simulaciones donde se incrementa la velocidad del flujo en la entrada de la fosa para observar el comportamiento de la formación del vapor en función de la velocidad del flujo (ver Fig. 7).
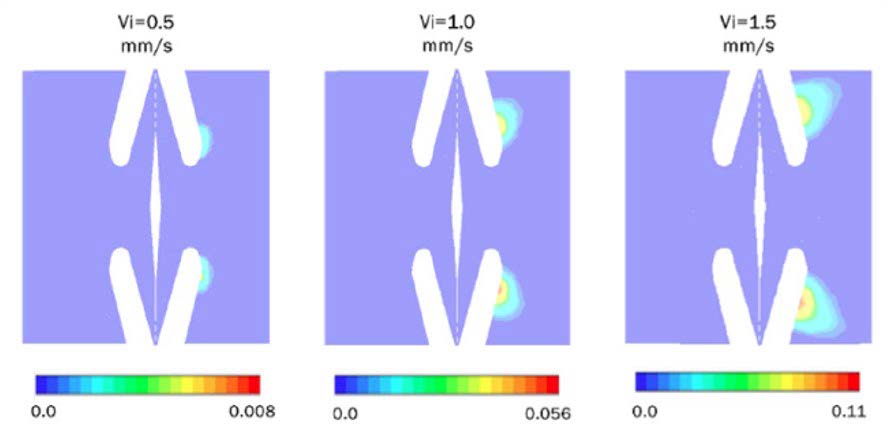
Figura 7. Formación de vapor como una función de velocidad de flujo en la entrada de la fosa. Se observan los contornos de la fracción volumétrica del vapor en función de la posición. La escala indica la fracción volumétrica en %
Conclusiones
Un ecosistema forestal saludable no es simplemente productivo, sino uno finamente equilibrado para la sostenibilidad de todas las especies que contribuyen a su existencia. Los árboles son el corazón de todo ecosistema y es difícil identificar algo más importante para la existencia de todos los seres vivos en la superficie de la Tierra. Actualmente, se favorece algunas especies basándose en consideraciones económicas; la ingeniería genética transforma a los favorecidos en clones "mejorados", pasando por alto la fisiología del árbol silvestre con la capacidad de supervivencia lograda por desafíos repetidos durante millones de años.
Recordemos que cada árbol no es sólo una biofábrica de producción de madera o frutos, sino un organismo extraordinario con una ascendencia que se remonta antes de la llegada del hombre
[10-11]. A partir de las simulaciones numéricas realizadas a diferentes casos, se obtienen los perfiles de presión entrada/salida de la fosa, así como los contornos de presión variando el número y tamaño de hoyos del margo de la fosa. Los resultados demuestran que el cambio en la diferencia de presión tiene una relación inversamente proporcional al incremento en el número y tamaño de los hoyos. Es decir, el flujo de agua dentro de los elementos del xilema pasa con mayor facilidad por aquellas fosas que tienen mayor cantidad de hoyos en su margo.
Además, con la visualización de los contornos de presión, se aprecia una zona de baja presión en la salida de la fosa, que eventualmente provoca la formación de vapor, que se extiende verticalmente hasta obstruir totalmente el conducto. Como trabajo futuro se planea realizar un análisis del flujo con condiciones saturadas de aire (nano burbujas), de tal manera, que sirvan como puntos de nucleación para una mayor generación de vapor, y eventualmente, encontrar la concentración para bloquear totalmente el conducto, comprobando así que el fenómeno de cavitación vaporosa sucede en las minúsculas fosas.
Agradecimientos
Trabajo patrocinado por el Programa de Apoyo a Proyectos de Investigación Tecnológico IN101712: Cavitación y embolia en árboles bajo cambio climático y contaminación ambiental.

Referencias
1] Dixon H. H. y Joly J. On the ascent of sap. Proceedings of the Royal Society of London 57(1894), 3-5.
-
Zimmermann M. H. Xylem structure and the ascent of sap, Berlin:
Springer Verlag, 1983. -
Brown H. R. The theory of the rise of sap in trees: some historical and conceptual remarks. Phys Perspect 15 (2013)320-39.
-
Brodribb T. J.; Bowman D. J.; M. S., Nichols S.; Delzon S. y Burlett R. Xylem function and growth rate interact to determine recovery rates after exposure to extreme water deficit. New Phytol 188(2010)533-10.
-
Zeppel M. J. B.; Macinnis-Ng CMO; Yunusa I. A.M.; Whitley R. J. y Eamus D. Long term trends of stand transpiration in a remnant forest during wet and dry years. J Hydrol 349(2008)200-14.
-
Vertessy R. A.; Watson F. G. y O'Sullivan R. S. K. Factors determining relations between stand age and catchment water balance in mountain ash forests. Ecol Manage 143(2002)13–26.
-
B. Zeppel M. J. B.; Anderegg W. R. L.; Adams H. D.; Hudson P.; Cook A.; Rumman R.; Eamus D.; Tissue D. T. y Pacala S. W. Embolism recovery strategies and nocturnal water loss across species influenced by biogeographic origin. Ecology and Evolution 9(2009) 5348-14.
-
Brunner I.; Herzog C. y Dawes A. D. Arend M., Sperisen C. How tree roots respond to drought. Front. Plant Sci. 6(2015) 547-16.
-
Valli A.; Koponen A.; Vesala T. y Timonen J. Simulations of water flow through bordered pits of conifer xylem.
-
Edwards D. 2003. Xylem in early tracheophytes. Plant Cell Environ.
26(1): 57–72. -
Savidge R. A. Learning from the past – the origin of wood. The forestry chronicle August 2008.
-
Schulte PJ, UG Hacke and AL Schoonmaker. 2015. Pit membrane structure is highly variable and accounts for a major resistance to water flow through tracheid pits in stems and roots of two boreal conifer species. New Phytologist 208:102-113.